El modelo de mosaico fluido de la estructura de las membranas celulares
ABSTRACTO
Un modelo de mosaico fluido se presenta para la organización bruto y la estructura de las proteínas y los lípidos de las membranas biológicas. El modelo es consistente con las restricciones impuestas por la termodinámica. En este modelo, las proteínas que son parte integral de la membrana son un conjunto heterogéneo de moléculas globulares, cada una dispuestas en una estructura anfipática, es decir, con los grupos iónicos y polares altamente sobresalen de la membrana en la fase acuosa, y los grupos no polares en gran parte sepultado en el interior hidrofóbico de la membrana. Estas moléculas globulares están parcialmente incrustadas en una matriz de fosfolípidos. La mayor parte de los fosfolípidos está organizada como una doble capa discontinua, fluida, aunque una pequeña fracción de los lípidos pueden interactuar específicamente con las proteínas de membrana. La estructura de mosaico fluido es por lo tanto, formalmente análogo a una solución orientada en dos dimensiones de las proteínas integrales (o lipoproteínas) en la bicapa de fosfolípidos solvente viscoso. Experimentos recientes con una gran variedad de techniqes y varios sistemas de membrana diferentes se describen, todo lo cual instigar consistente con, y añadir mayor cantidad de detalles que, el modelo de mosaico fluido. Por lo tanto, parece apropiado sugerir posibles mecanismos para funciones de la membrana y fenómenos mediados por membrana a la luz del modelo. A modo de ejemplo, los mecanismos comprobables experimentalmente, se sugieren los cambios de la superficie celular en la transformación maligna, y para efectos de cooperación expuesto en las interacciones de las membranas con algunos ligandos específicos.
Nota añadido en la prueba: Dado que este artículo fue escrito, se han obtenido pruebas de microscopía electrónica (69) que la concanavalina A los sitios de unión en las membranas de fibroblastos de ratón transformado SV40-virus (células 3T3) están más agrupadas que los sitios en las membranas de las células normales, como predice la hipótesis representada en la figura. 7B. T-aquí también ha aparecido un estudio de Taylor et al. (70) que muestra los efectos notables producidos en los linfocitos de la adición de anticuerpos dirigidos a las moléculas de inmunoglobulina de superficie. Los anticuerpos induce una redistribución y pinocitosis de estas inmunoglobulinas de superficie, de manera que en unos 30 minutos a 37 ° C las inmunoglobulinas de superficie están completamente barridos de la membrana. Estos efectos no se producen, sin embargo, si los anticuerpos bivalentes se sustituyen por los fragmentos Fab monovalente o si los experimentos de anticuerpos se llevan a cabo a 0 ° C en lugar de 37 º C. Resultados de estos y otros indican claramente que los anticuerpos bivalentes produce una agregación de las moléculas de inmunoglobulina de superficie en el plano de la membrana, que puede ocurrir sólo si las moléculas de inmunoglobulina son libres de difundir en la membrana. Esta agregación se parece desencadenar la pinocitosis de los componentes de la membrana por algún mecanismo desconocido. Tales transformaciones membrana puede ser de crucial importancia en la inducción de una respuesta de anticuerpos a un antígeno, así como los procesos iv otras de la diferenciación celular.
Las membranas biológicas juegan un papel crucial en casi todos los fenómenos celulares, aun nuestro entendimiento de la organización de las membranas es rudimentaria. Nosotros sugerimos que existe una analogía entre los problemas de la estructura de la membrana y de las proteínas.
Singer ha examinado recientemente en muchos modelos detallados considerables de la organización estructural grosa de las membranas, en términos de la termodinámica de los sistemas macromoleculares y en la luz de la entonces evidencia experimental disponible. De este análisis, fue concluido que la estructura de mosaico de las proteínas globulares alternantes y bicapa de fosfolípidos fue el único modelo de membrana entre aquellos analizados que fue simultáneamente consistente con las restricciones termodinámicas en que todos los datos experimentales.
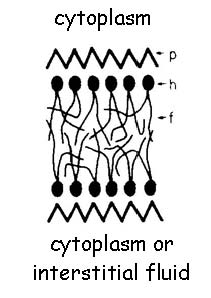
Figura 1: la figura original de Singer (1992) ilustra el modelo de Davson-Danielli-Robertson de la membrana plasmática. Tenga en cuenta que la bicapa lipídica se aísla de su entorno acuoso por dos capas de proteína de la membrana se desarrolló (p). Cada membrana de lípidos que forman está compuesta por un grupo de cabeza polar (h) y la cola acilo graso (f)
Desde que el artículo fue escrito, mucha nueva evidencia ha sido publicada y apoya fuertemente el modelo de mosaico. En particular el mosaico parece ser fluida y dinámica para muchos propósitos. En este artículo por tanto presentamos y discutimos un modelo de mosaico fluido de estructura de membrana y proponemos que es aplicable a la mayoría de membranas biológicas tales como plásmalema y membranas de diferentes organelos celulares tales como mitocondria y cloroplastos.
El modelo de mosaico fluido ha evolucionado por una serié de estadios de versiones previas o anteriores. Las consideraciones termodinámicas acerca de las membranas y componentes de membranas iniciados, son aun centrales a estos desarrollos a esos estadios.
Hay otras interacciones no covalentes, tal como la unión a hidrogeno en interacciones electrostáticas, que también contribuyen a diferentes estructuras macromoléculas, sin embargo con respecto a la estructura grosa hay muchas probabilidades de magnitud secundaria comparado con interacciones hidrófobicas e hidrofilicas.

Figura 2: la figura original de Singer y Nicolson (1972) que representa la sección transversal de membrana con las proteínas integrales en el mosaico bicapa de fosfolípidos. Los fosfolípidos son representados como esferas con las colas, las proteínas como objetos incrustados sombra, globular. Proteínas periféricas, que se situaría en, no en la superficie de la membrana, no se muestran. Hay que recordar que tanto las superficies de esta membrana interceptar un medio acuoso o en el citoplasma y / o el líquido intersticial. Proteína transmembrana que abarcan la membrana entera a la izquierda.
La bicapa fosfolípida ilustro los efectos combinados de interacciones hidrofobicas e hidrofilicas en esta estructura las cadenas de ácidos grasos no polares de los fosfolípidos son secuestrados juntos lejos del contacto con agua por tanto maximiza la interacciones hidrofobicas más aun los grupos iónicos y bipolares están en contacto directo con la fase acuosa en la superficie exterior de la bicapa , por tanto hidrofilicas . En el caso de fosfolípidos bipolares tales como fosfatidil colina, las interacciones bipolares entre los pares de iones en la superficie de la bicapa pueden contribuir también a la estabilidad de la estructura de la bicapa.

Figura 3: la figura original de Singer y Nicolson (1972) que representa la sección transversal con las proteínas de membrana integral en la bicapa de fosfolípidos. Las porciones iónicas y polares de las proteínas, como lo indica el + / - signos, póngase en contacto con las soluciones acuosas (citoplasma y / o líquido intersticial) que rodea a la bicapa lipídica. La membrana que abarca o se inserta la región de la proteína es non-polar/hydrophobic y por lo tanto carece de carga, como se indica por la ausencia de + / - símbolos.
Bibliografía:
Science 18 February 1972:
Vol. 175 no. 4023 pp. 720-731
DOI: 10.1126/science.175.4023.720
Vol. 175 no. 4023 pp. 720-731
DOI: 10.1126/science.175.4023.720
The Fluid Mosaic Model of the Structure of Cell Membranes
1. S. J. Singer and
No hay comentarios:
Publicar un comentario